Of the approximately 10,000 species of birds in the world, 6000 of them belong to a single order, the Passeriformes, compared with two species of rhea (order Rheiformes) or just a single hoatzin (order Opisthocomiformes). What has caused this asymmetry?
Why do some sets of closely-related birds look and act essentially the same (say, Phylloscopus warblers) whereas others have vastly different morphologies (e.g., ovenbirds) and behaviours (e.g., manakins, birds of paradise)?
And once we have answers to this -- are birds particularly special in this regard? What can other taxa tell us about general evolutionary principles underlying the diversity of life on earth?
The best way to find out more about my and my students' recent work is via my publications and pre-prints listed on Google Scholar, but I've summarised some recent and ongoing research threads below.
Why do some sets of closely-related birds look and act essentially the same (say, Phylloscopus warblers) whereas others have vastly different morphologies (e.g., ovenbirds) and behaviours (e.g., manakins, birds of paradise)?
And once we have answers to this -- are birds particularly special in this regard? What can other taxa tell us about general evolutionary principles underlying the diversity of life on earth?
The best way to find out more about my and my students' recent work is via my publications and pre-prints listed on Google Scholar, but I've summarised some recent and ongoing research threads below.
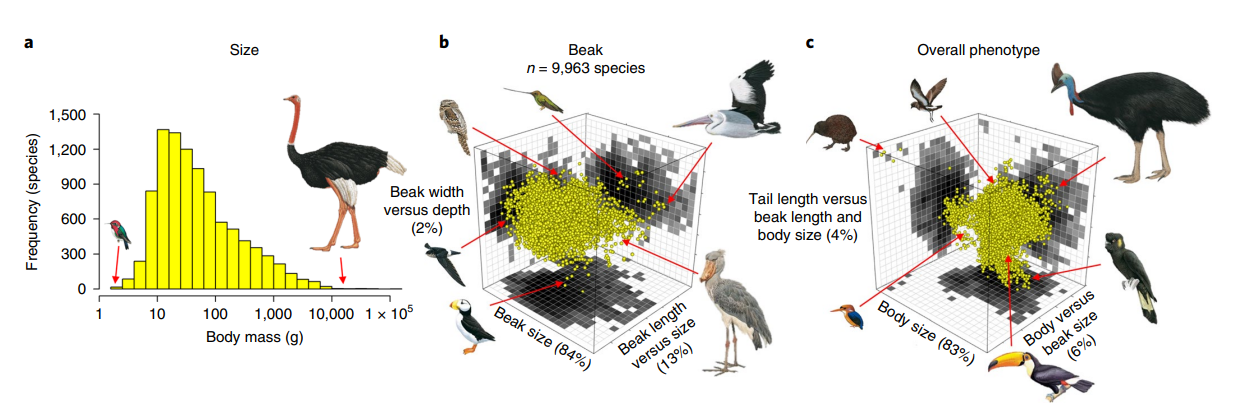
Image from Pigot*, Sheard*, et al. 2020, Nature Ecology & Evolution
The Evolution of Morphology
How do we quantify a species' role in an ecosytem? In a way that can be both standardized across large numbers of species and feasibly collected on such a sample? One way is to use museum specimens (and, where possible, live-caught species) to measure morphological traits, quantifying species' size and shape.
In what Science Magazine called an "ecological gold mine", I was the single largest contributor to, and one of the early coordinators of, AVONET, a database of functional traits for the world's birds. I have then used (and am still using) this data to explore broad-scale avian morphological evolution. For example, we can use a machine learning technique called random forest modelling to then link these beak measurements to global variation in diet and foraging strategy (with decent success, though these sorts of relationships tend to break down at smaller taxonomic scales); we can also use wing measurements as a proxy for flight ability and employ phylogenetic mixed models to examine the relationship between flight, behaviour, and the environment.
I am currently working on building similar large-scale morphological databases in mammals and spiders.
In what Science Magazine called an "ecological gold mine", I was the single largest contributor to, and one of the early coordinators of, AVONET, a database of functional traits for the world's birds. I have then used (and am still using) this data to explore broad-scale avian morphological evolution. For example, we can use a machine learning technique called random forest modelling to then link these beak measurements to global variation in diet and foraging strategy (with decent success, though these sorts of relationships tend to break down at smaller taxonomic scales); we can also use wing measurements as a proxy for flight ability and employ phylogenetic mixed models to examine the relationship between flight, behaviour, and the environment.
I am currently working on building similar large-scale morphological databases in mammals and spiders.
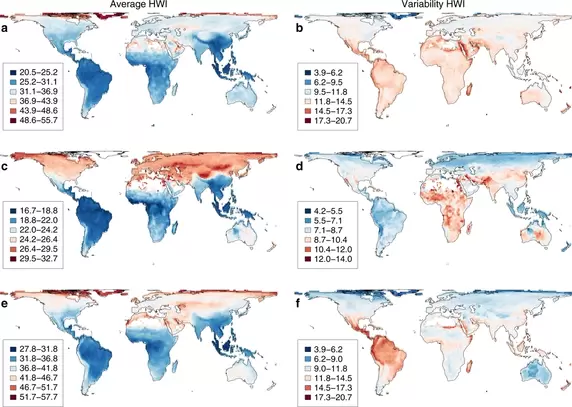
A map of variation in avian wing shape, from Sheard et al. 2020 Nature Communications
Causes and Consequences of Parental Care
The presence and extent of parental care can influence not only an individual's reproductive success, but can also affect the macroevolutionary trajectory of the entire lineage. I am interested in not only what causes variation in care behaviours, but also how these care behaviours then affect other traits and what the macroevolutionary consequences are of these species-wide reproductive investments.
One stunning example of parental care behaviours in the animal kingdom is the construction of the avian nest. There have been some surprising results when studying this trait at the global scale. For example, contrary to popular wisdom, bird nests do not correlate with variation in egg shape at the global scale (click here for some fun data viz about this project, or see coverage of this study in the New York Times or the Atlantic, or catch us mentioned by David Attenborough), and nest structure mostly doesn't correlate with chick development. Nest material use and beak shape, however, do correlate, as does inter-specific variation in nest size and temperature.
I've also -- with my former PhD student Karina Vanadzina -- worked on parental care in fish.
One stunning example of parental care behaviours in the animal kingdom is the construction of the avian nest. There have been some surprising results when studying this trait at the global scale. For example, contrary to popular wisdom, bird nests do not correlate with variation in egg shape at the global scale (click here for some fun data viz about this project, or see coverage of this study in the New York Times or the Atlantic, or catch us mentioned by David Attenborough), and nest structure mostly doesn't correlate with chick development. Nest material use and beak shape, however, do correlate, as does inter-specific variation in nest size and temperature.
I've also -- with my former PhD student Karina Vanadzina -- worked on parental care in fish.

A giant panda, an excellent example of a black-and-white mammal. (Photo by Catherine Sheard.)
The Evolution of Colour
At the moment, I'm thinking a lot about the macroevolutionary causes and consequences of animal colour. Mostly, this is watch-this-space, but in the meantime you can go read the work of some of my former students, such as Natasha Howell's excellent paper on the origins of black-and-white colouration in mammals or Rob MacDonald's work on the link (or, rather, lack thereof) between primate colouration and colour vision!
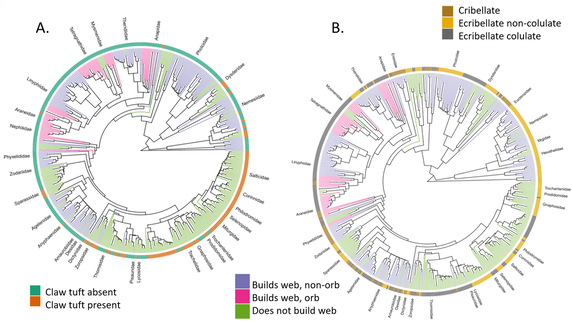
Morphological and behavioural traits plotted on spider phylogenies, from a project currently under revision
Phylogenetic scale
Finally, I am currently thinking a lot about the mathematical, ecological, and evolutionary underpinnings of a phenomenon sometimes known as "phylogenetic scale", where one set of macroevolutionary results can be found at one hierarchical level (say, a class or an order) but not at another (say, an order or family). Cassie Stoddard and I (and a great team of collaborators) wrote about the phenomenon of phylogenetic scale in the evolution of egg shape here, but it's cropping up again and again across the field of trait macroevolution, in my work and in others'. This isn't even a "watch this space" as much as a PSA -- if you do this type of broad-scale comparative research but aren't also thinking hard about phylogenetic scale, maybe you should!
